Modelli di evoluzione di una popolazione isolata
Il calcolo della crescita di una popolazione è stato spesso oggetto di studio nel corso del tempo, sia dal punto di vista biologico che matematico. Analizzando diverse popolazioni o confrontando le possibili relazioni di crescita che possono legare due specie, sono stati ottenuti diversi modelli matematici validi, spesso basati sull’uso di logaritmi e funzioni esponenziali.
Per studiare l’evoluzione di una popolazione nel tempo bisogna utilizzare un modello dinamico, cioè un modello che tenga conto delle variazioni che avvengono in un sistema al trascorre del tempo, in cui il valore della grandezza ad un certo tempo t (variabile in ingresso) influenza il valore della grandezza al tempo successivo (variabile in uscita). Dal punto di vista matematico un sistema dinamico può essere descritto da un processo iterativo, cioè da un processo che consiste nell'applicare ripetutamente una stessa trasformazione.
Se con T si indica una generica trasformazione che in corrispondenza ad un input i produce un output u=T(i), partendo da uno start i0, per effetto della trasformazione T si ottiene un’uscita u0; se si immette come nuovo ingresso, il valore u0, cioè i1=u0 si ottiene in corrispondenza un’uscita u1, (questa operazione è detta di retroazione o feed-back). Così procedendo, iterativamente, si realizza un processo che all’n-esimo passo ha come ingresso l’uscita del passo precedente (n-1) e l'uscita costituirà l’ingresso del passo successivo.
Il processo è descritto formalmente dal seguente sistema:
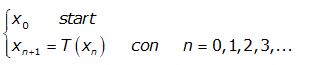
dove xn è detta successione delle iterate.
Di seguito sono illustrati alcuni modelli della dinamica dell’evoluzione della specie studiati sin dagli inizi del secolo XlX.
Crescita della popolazione e crescita delle risorse secondo la teoria maltusiana
Thomas Robert Malthus e il Malthusianesimo
 Thomas R. Malthus (Londra 1766 – Haileybury 1834), figlio di un giacobino, fu educato privatamente sino a diciotto anni e proseguì gli studi al Jesus college di Cambridge dove conseguì ottimi risultati nonostante avesse un marcato difetto di parola. Dal 1805 fu professore di Economia Politica presso il college di Haileybury, dove venivano educati i funzionari dell’East India Company.
Thomas R. Malthus (Londra 1766 – Haileybury 1834), figlio di un giacobino, fu educato privatamente sino a diciotto anni e proseguì gli studi al Jesus college di Cambridge dove conseguì ottimi risultati nonostante avesse un marcato difetto di parola. Dal 1805 fu professore di Economia Politica presso il college di Haileybury, dove venivano educati i funzionari dell’East India Company.
Mentre si trovava a Cambridge, prese i voti diventando curato della Chiesa di Inghilterra e nel 1796 divenne parroco di Albury. Si sposò a 38 anni ed ebbe tre figli. Nel 1798 pubblicò il volume Essay on the Principle of Population (Saggio sul principio di popolazione), che è citato dallo stesso Darwin nella sua autobiografia. Il saggio descrive la teoria economica da lui ipotizzata chiamata appunto Malthusianesimo; esso studia il rapporto esistente fra popolazione e risorse naturali disponibili sul nostro pianeta.
Saggio sul principio di popolazione
La prima edizione venne pubblicata in anonimo nel 1798, mentre la seconda, accresciuta e modificata, venne ristampata nel 1803. Il punto di partenza è l'analisi del rapporto tra l’accrescimento della popolazione e l’accrescimento dei mezzi di sussistenza. Malthus osserva che la popolazione tende a crescere secondo una progressione geometrica (2, 4, 8, …), con tempo di raddoppiamento di venticinque anni, mentre i mezzi di sussistenza tendono a crescere secondo una progressione aritmetica (1, 2, 3, …). Lo squilibrio che così si determina tra popolazione e mezzi di sussistenza può essere, secondo Malthus, eliminato in primo luogo attraverso la naturale funzione di “controllo repressivo” esercitata dalla miseria e dal vizio, che riducono la popolazione; in secondo luogo attraverso il controllo preventivo delle nascite. Sarebbe auspicabile che il progresso della società umana consistesse nel sostituire il più possibile il controllo repressivo con il controllo preventivo: ad esempio impedendo l’accrescersi eccessivo della popolazione mediante quello che l’economista chiama il «ritegno morale», cioè «con l’astenersi dal matrimonio per motivi prudenziali e con una condotta strettamente morale durante il periodo di questa astinenza».
Il seguente grafico rappresenta un esempio di crescita di popolazione in relazione alla crescita delle risorse.
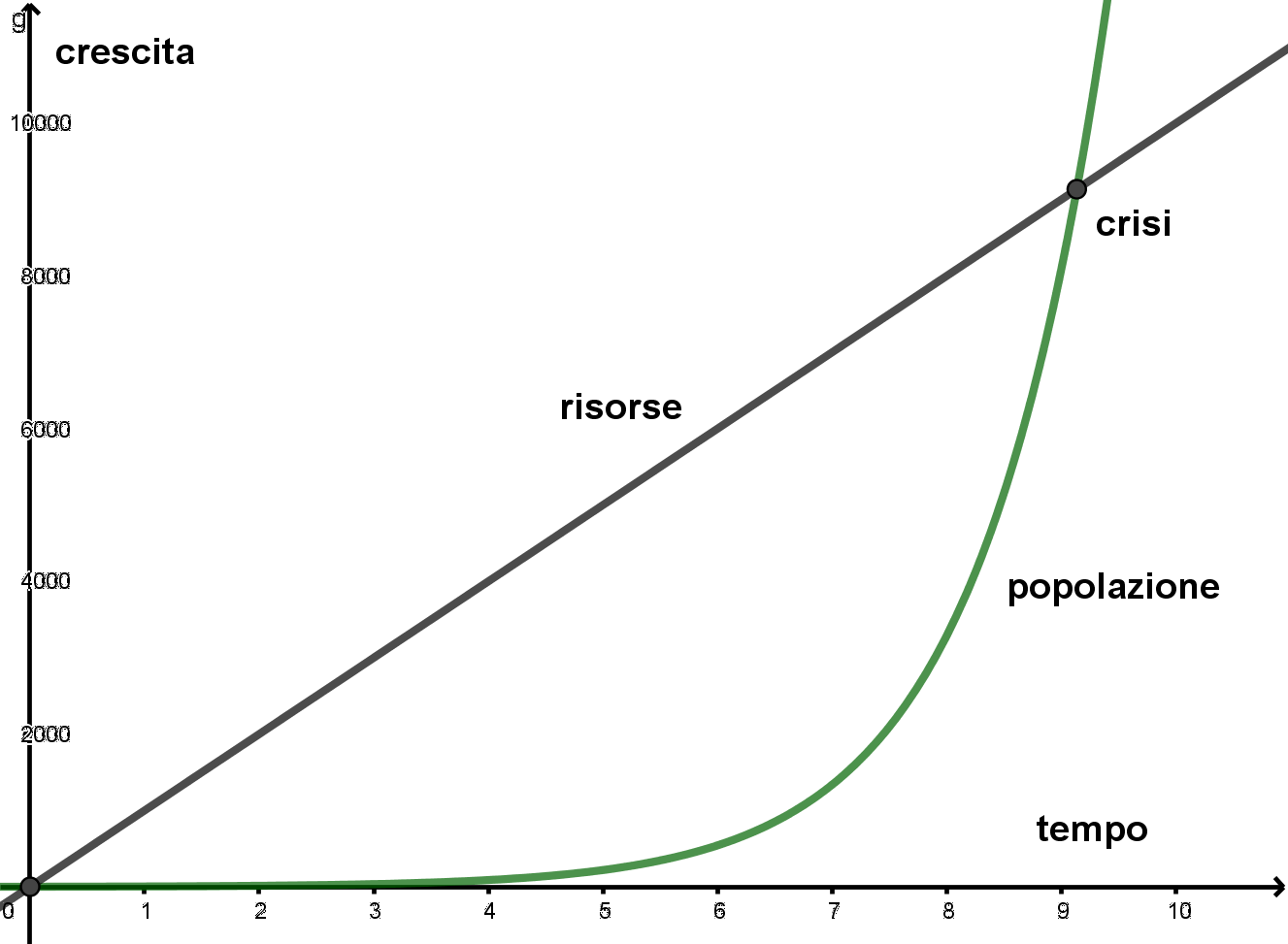
È pur vero che il modello malthusiano si adatta ad un'economia preindustriale nella quale il tasso di crescita delle risorse non tiene conto di una produttività migliorata attraverso il progresso tecnologico.
Modello matematico
Malthus per studiare una popolazione isolata, cioè una popolazione formata da individui che convivono senza scambi con l’esterno né in termini di risorse né in termini di individui, ha proposto il seguente modello iterativo:
se si indica con xn il numero di individui presenti allo stadio n, o tempo t, la variazione di popolazione fra due stadi consecutivi è data dalla differenza
![]()
che prende il nome di tasso di crescita. Il tasso di crescita può essere positivo (la popolazione cresce), negativo (la popolazione decresce) o nullo (il numero di individui rimane costante).
Malthus ipotizza che il tasso di crescita sia direttamente proporzionale al numero di individui:
![]()
dove a è fattore di crescita della popolazione.
Il processo iterativo che descrive l’evoluzione della popolazione è
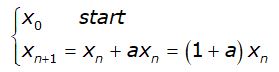
ed è espresso dalla funzione lineare
![]()
Se si ripete il processo n volte si ottiene
![]()
che rappresenta il termine generale di una progressione geometrica, di ragione (1+a). L’evoluzione del processo dipende dal valore di a, se a=0 il processo è stazionario, la crescita della popolazione è nulla; se a>0 il processo è in esplosione, la crescita della popolazione è in espansione; se a<0 il processo è in estinzione, la popolazione è in estinzione.
Dal modello iterativo descritto, considerato il legame tra la progressione geometrica e la funzione esponenziale esposto nel capitolo precedente, quando da un insieme discreto si passa ad un insieme continuo si ottiene la legge di evoluzione della popolazione secondo Malthus:
![]()
con
- N(t) la popolazione al tempo t,
- N0 la popolazione al tempo zero,
- g differenza tra il tasso di natalità ed il tasso di mortalità.
Al variare del valore di g è possibile prevedere l’andamento della popolazione:
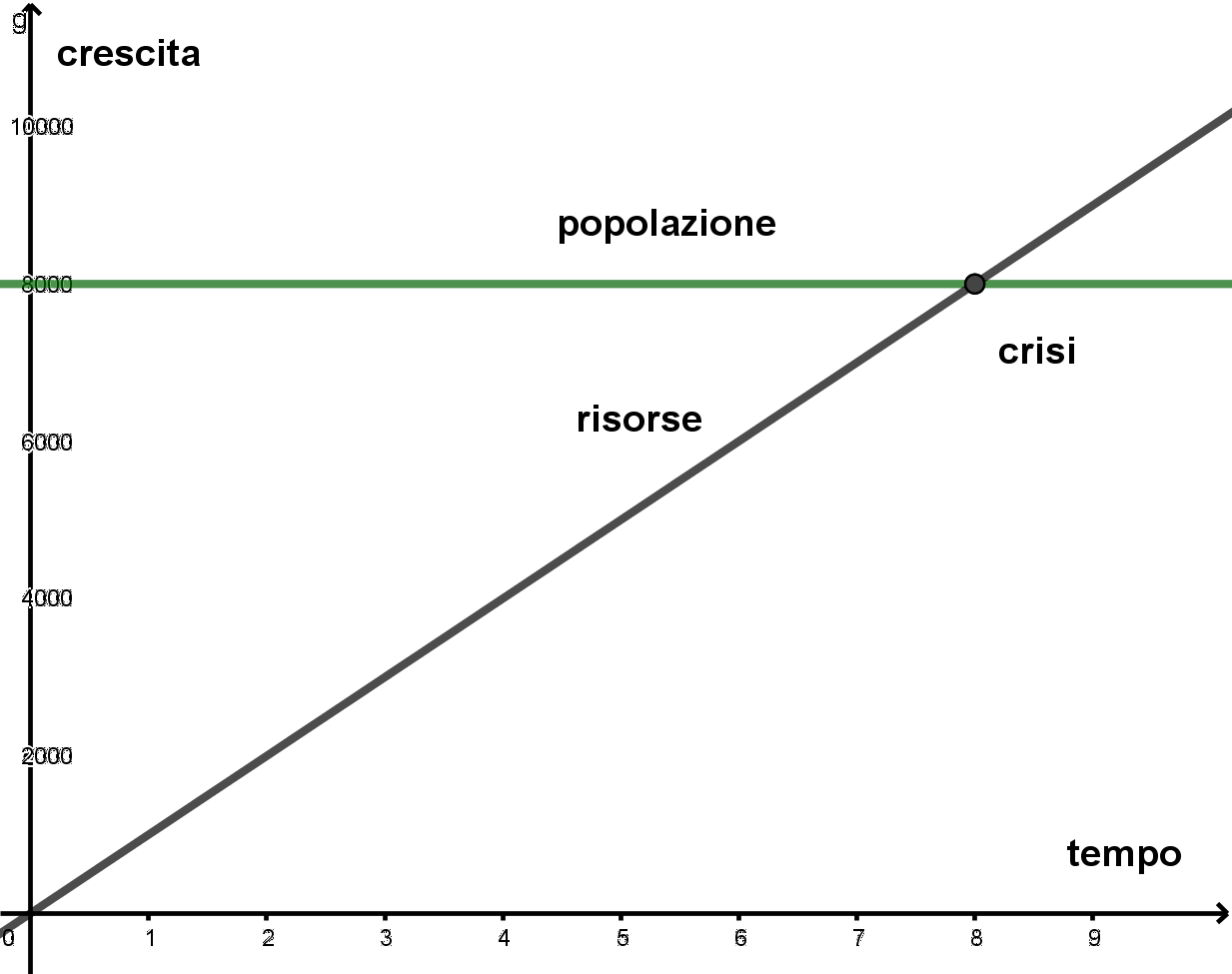
- Per g=0 l’equazione di Malthus diventa N(t)=N0e0t=N0, quindi la popolazione è quindi rimasta invariata. Il grafico che segue rappresenta la crescita di una popolazione per g=0
- Per g>0 la popolazione cresce in modo esponenziale, il numero di nati supera il numero di morti. Il grafico che segue rappresenta la crescita di una popolazione per g>0
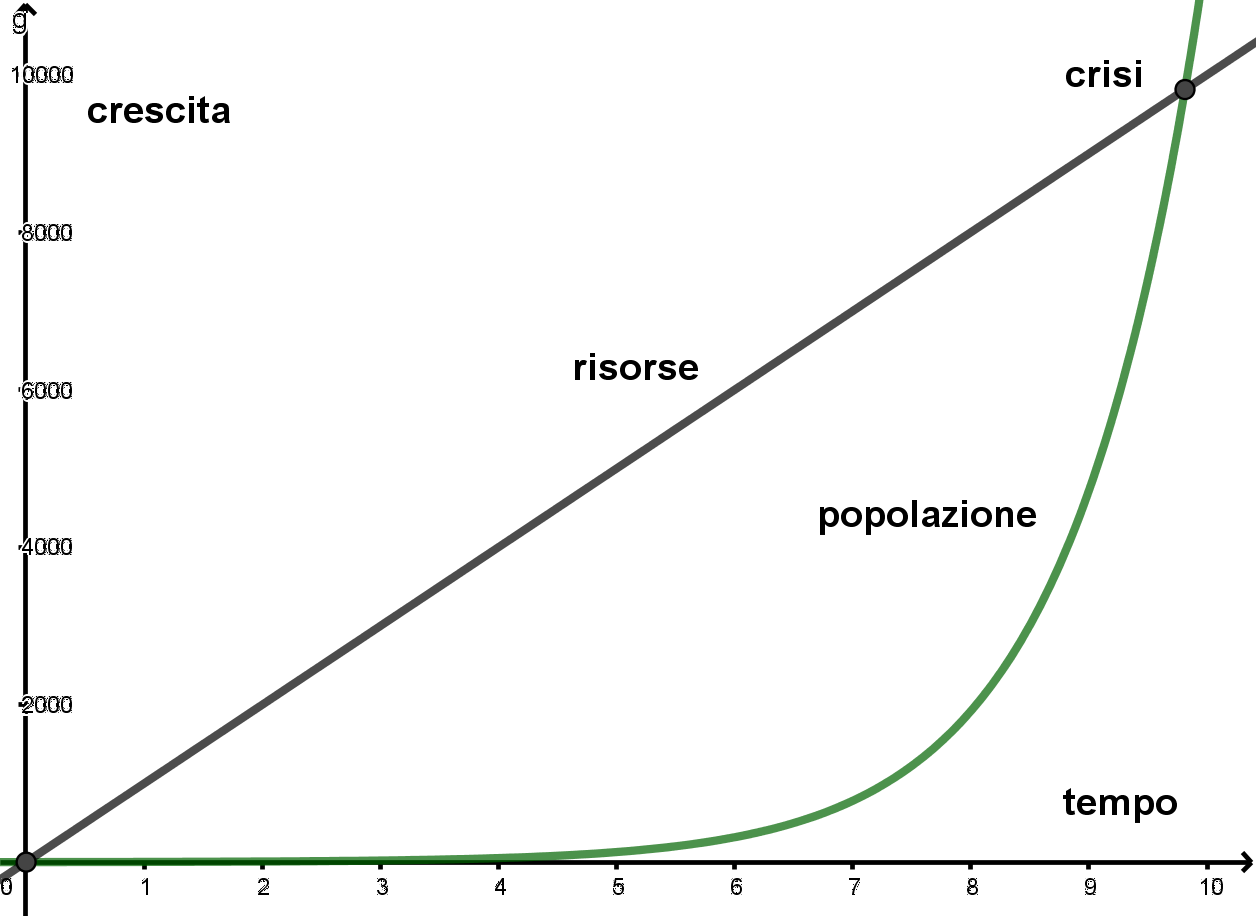
- Per g<0 la popolazione decresce in modo esponenziale ed è quindi destinata all’estinzione. Il grafico che segue rappresenta la crescita di una popolazione per g<0
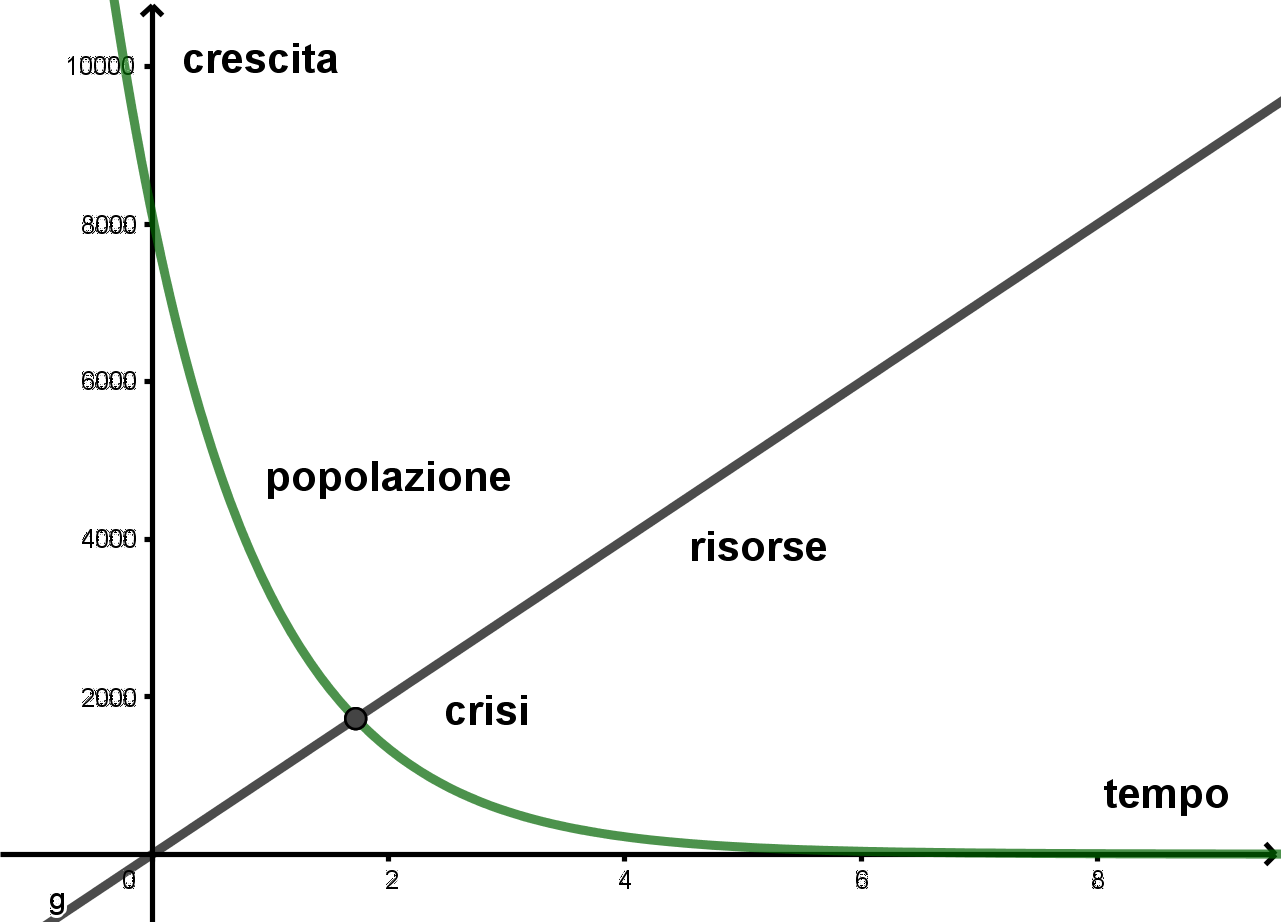
Dalla legge di evoluzione della popolazione secondo Malthus
![]()
è possibile, grazie all’uso dei logaritmi, determinare il tempo di evoluzione ed il tasso di crescita di una popolazione.
Esempio 1
La seguente tabella riporta le proiezioni delle 10 nazioni più popolose della popolazione mondiale.
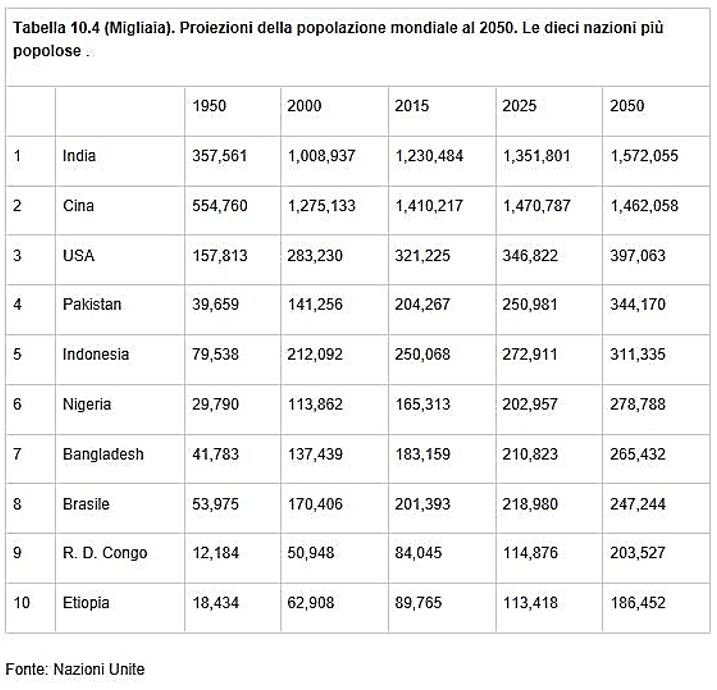
Adottando il modello di crescita di Malthus, si può determinare il fattore di crescita della popolazione di una nazione, ad esempio, la Repubblica Del Congo nell’intervallo di tempo compreso tra il 2015 e 2050.
Dalla tabella si considerano i seguenti valori
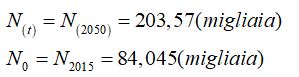
Il fattore di crescita della popolazione risulta quindi:
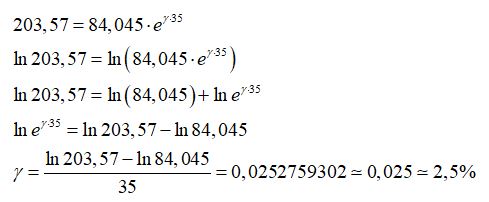
Il fattore di crescita della popolazione della Repubblica Del Congo dal 2015 al 2050 è pari 2,5% ed è quindi una popolazione destinata ad “esplodere” secondo il modello di Malthus.
Esempio 2
L’ASviS (Alleanza Italiana per lo Sviluppo Sostenibile) il 27 giugno 2017, ha pubblicato un articolo a cura di Lucilla Persichetti dal titolo “L’Onu aumenta le sue stime per la popolazione mondiale: 9,8 miliardi nel 2050”
Con il World Population Prospects (Prospetto della popolazione mondiale) 2017 i pronostici per la popolazione al 2050 aumentano di 100 milioni di individui rispetto alle ultime previsioni. Tra le cause principali la rapida crescita demografica in Africa e India.
Il World Population Prospects viene elaborato ogni due anni dalla "Divisione per la popolazione del Dipartimento per gli affari economici e sociali delle Nazioni Unite" e fornisce stime e proiezioni relative alla situazione demografica mondiale.
La tabella che segue riporta i dati della crescita della popolazione riportati nel Prospetto della popolazione mondiale
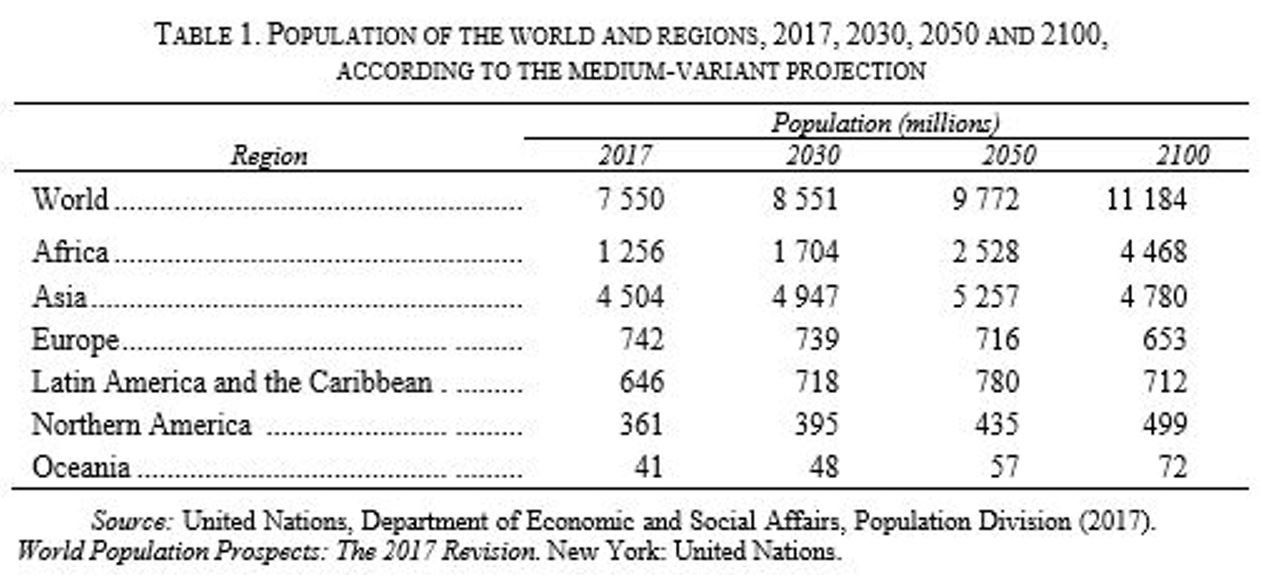
Adottando il modello di crescita di Malthus determinare
- il fattore di crescita della popolazione dell’Africa e dell’Europa nell’intervallo di tempo compreso tra il 2017 e 2030;
- l’anno in cui l’Europa raggiungerà una popolazione pari a 600 milioni, nell’ipotesi che il tasso di crescita della popolazione sia negativo, del -0,03% e considerando come popolazione al tempo zero quella del 2017.
Punto a
In riferimento alla popolazione dell’Africa si ha
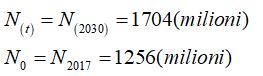
il fattore di crescita della popolazione è
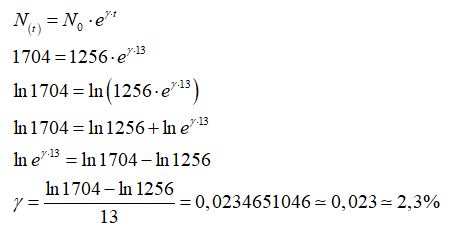
Il fattore di crescita della popolazione dell’Africa dal 2017 al 2030 è pari 2,3% ed è quindi una popolazione destinata ad “esplodere” secondo il modello di Malthus.
In riferimento alla popolazione dell’Europa si ha
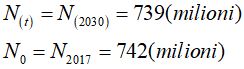
il fattore di crescita della popolazione è
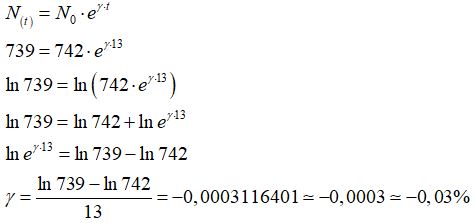
Il fattore di crescita della popolazione dell’Europa dal 2017 al 2030 è pari -0,03%.
Punto b
Considerati i seguenti dati
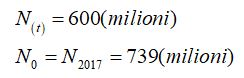
si ha:
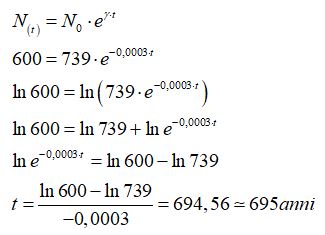
Nell’ipotesi che il tasso di crescita della popolazione sia negativo del -0,03% e considerando come popolazione al tempo zero quella del 2017, secondo il modello di Malthus, la popolazione dell’Europa sarà pari a 600 milioni nell’anno 2712.
Modelli di iterazione tra due popolazioni
Il modello teorico
L’analisi della dinamica di coesistenza di più specie nello stesso ambiente risulta complessa rispetto allo studio dell’evoluzione di una singola, anche nel caso in cui le risorse siano illimitate e l’habitat chiuso.
Il primo ad occuparsi di ricerche riguardati le relazioni tra due specie fu il matematico italiano Vito Volterra (Ancona il 3 maggio 1860, Roma l'11 ottobre 1940), il quale nel 1926 elaborò un modello matematico riguardante la dinamica 'preda-predatore', detto il modello lineare di Volterra.
Indicati con xn ed yn il numero di individui di ciascuna specie presenti allo stadio n, tale modello suppone che il tasso di crescita delle due popolazioni sia direttamente proporzionale al numero degli individui di ciascuna, con fattore di crescita costante ed è espresso dalle seguenti relazioni
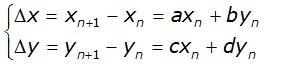
dove a e d sono i coefficienti di crescita di ciascuna popolazione rispetto a se stessa, b e c di ciascuna rispetto all'altra. Ricavando xn+1 e yn+1 il processo iterativo che descrive l'evoluzione delle due popolazione è
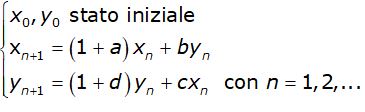
Ponendo
![]()
si ottiene il modello
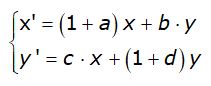
che rappresenta un’estensione del modello di Malthus al caso di due popolazioni.
Nella tabella che segue sono rappresentate alcune possibili evoluzioni di due popolazioni al variare del segno (positivo o negativo) dei coefficienti di crescita di ciascuna delle due popolazioni:
|
Segno dei coefficienti di crescita |
Evoluzione dell’eco-sistema |
|||
|
a |
b |
c |
d |
|
|
- |
+ |
+ |
- |
COOPERAZIONE |
|
+ |
- |
- |
+ |
COMPETIZIONE |
|
+ |
- |
+ |
- |
PREDA-PREDATORE |
Regime di cooperazione
In tale regime:
- i coefficienti di crescita a e d (di ciascuna popolazione rispetto sé stessa) sono negativi e pertanto, secondo il modello malthusiano, ciascuna popolazione, considerata singolarmente si estinguerebbe;
- i coefficienti di crescita b e c invece (di ciascuna popolazione rispetto all’altra) sono positivi e di conseguenza la presenza dell’altra specie può rallentare o interrompere il processo di estinzione.
La cooperazione permette alle due specie di evolvere verso uno stato di equilibrio.
Regime di competizione
In tale regime:
- i coefficienti di crescita a e d (di ciascuna popolazione rispetto sé stessa) sono positivi,
- i coefficienti di crescita b e c (di ciascuna popolazione rispetto all’altra) sono negativi,
pertanto, ciascuna specie in assenza dell'altra esploderebbe.
La competizione fa sì che una delle due specie si estingue e l’altra esplode.
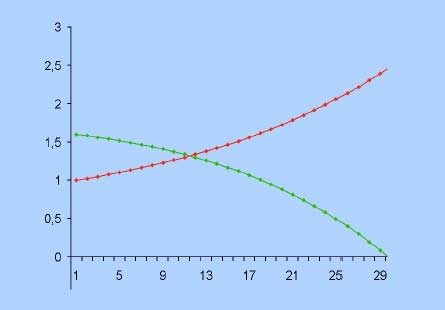
Regime preda-predatore
In tale regime:
- i coefficienti di crescita della prima specie a e c sono positivi
- i coefficienti di crescita della seconda specie b e d sono negativi
La popolazione y dei predatori inibisce la crescita della popolazione preda x che da sola esploderebbe. A sua volta la popolazione x invece evita il collasso al quale sarebbe destinata la popolazione y se fosse sola. I predatori sopravvivono grazie alle prede, le quali crescerebbero in maniera illimitata se non fossero tenute sotto controllo dai primi. Nella loro reciproca iterazione i predatori hanno la meglio sulle prede che dopo un certo tempo si estinguono, provocando la successiva estinzione dei predatori.
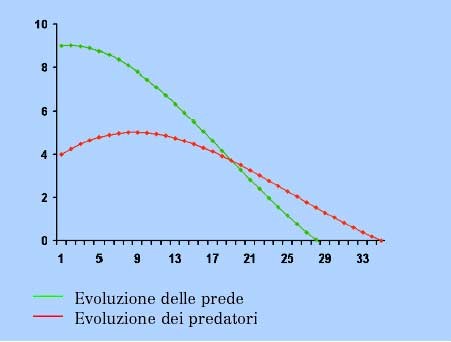
Le immagini riportate sono state prese dall’articolo “La modellizzazione del quotidiano come motore di innovazione didattica” del gruppo ricerca PRISTEM http://matematica-old.unibocconi.it/brandi/modelli2.htm
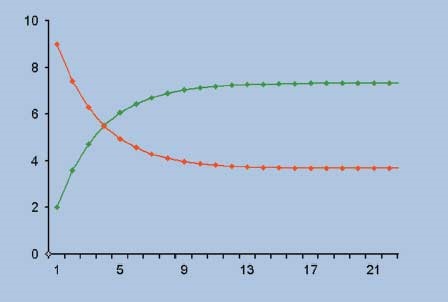
Modello preda-predatore non lineare di Lotka-Volterra
Vito Volterra per lo studio del regime preda-predatore, ha proposto il seguente modello
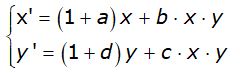
che è un’evoluzione non-lineare del modello precedente. Il termine xy rappresenta il numero di incontri tra prede e predatori e i coefficienti b e c sono le percentuali dell'esito di tali incontri che portano alla morte delle prede.
Una rappresentazione grafica del modello di tipo non lineare può essere quello riportato nella figura seguente, secondo il quale il modello del bio-sistema preda-predatore può produrre un comportamento oscillante non periodico, suddiviso in quattro fasi.
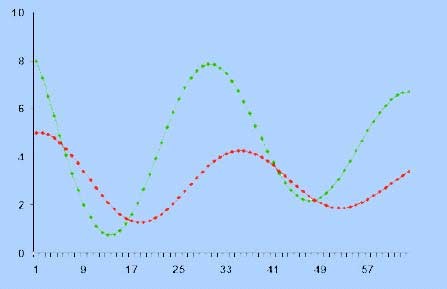
il grafico rosso rappresenta le prede, il grafico verde rappresenta i predatori
- Fase 1: i predatori crescono e si riproducono, a spese delle prede che diminuiscono progressivamente.
- Fase 2: le prede non sono sufficienti a soddisfare le eccessive esigenze dei predatori che cominciano a morire.
- Fase 3: a mano a mano i predatori diminuiscono le prede aumentano di numero.
- Fase 4: la popolazione dei predatori, avendo più cibo a disposizione, si riprende e il ciclo ricomincia.
Un esempio del regime preda-predatore: volpi e conigli
Volpi e conigli, ovvero predatori e prede, che convivono in uno stesso habitat, sono fra loro in perenne conflitto e insieme perennemente alleati per sopravvivere.
Se le volpi uccidono troppi conigli, questi rischiano di estinguersi, ma allora anche le volpi, rimaste senza cibo, li seguiranno nella stessa sorte. Il calo del numero di predatori, però, può salvare i conigli dall'estinzione. Tuttavia, se i conigli uccisi sono troppo pochi, il loro numero rischierà di crescere a dismisura e le risorse ambientali potranno diventare insufficienti. Anche se le risorse fossero illimitate, le due popolazioni non crescerebbero indefinitamente: la maggior disponibilità di cibo farebbe aumentare anche le volpi, ma in tal caso anche la predazione aumenterebbe e la crescita della popolazione dei conigli subirebbe un rallentamento, se non addirittura un'inversione.
Il modello grafico
 Il processo può essere rappresentato anche come una curva chiusa che si va allargando col passare del tempo: sembra un ciclo che si ripete, con i conigli e le volpi che rischiano sempre di estinguersi, ma poi si salvano e alla lunga diventano sempre più numerosi.
Il processo può essere rappresentato anche come una curva chiusa che si va allargando col passare del tempo: sembra un ciclo che si ripete, con i conigli e le volpi che rischiano sempre di estinguersi, ma poi si salvano e alla lunga diventano sempre più numerosi.
Il punto centrale corrisponde al minimo della funzione H(conigli,volpi) ed è un punto di equilibrio stabile: il sistema oscilla in continuazione su varie curve di livello, ma poi torna sempre in quel punto particolare.
Il grafico qui accanto mostra come variano prede (asse delle ascisse) e predatori (ordinate) nel tempo: il sistema inizia la sua evoluzione con le prede che aumentano notevolmente, corrispondente a un aumento, minore, dei predatori; quando, però, le prede sono in numero molto alto, i predatori iniziano ad aumentare di gran lunga e, di conseguenza, le prede diminuiscono fino a quando non saranno così poche che anche i predatori inizieranno a diminuire: in questo periodo, sia prede che predatori diminuiranno, tornando allo stato iniziale del sistema.
L'evoluzione prede-predatori è tutt'altro che scontata: ci sono due periodi in cui diminuiscono e aumentano entrambi, mentre, in altri due tratti, al diminuire di uno aumenta l'altro.
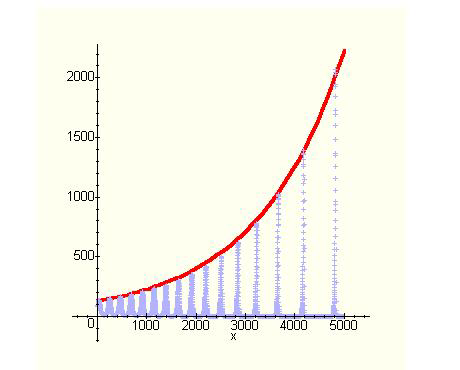 La crescita delle popolazioni avviene comunque in maniera esponenziale, nel senso che la sequenza dei massimi della popolazione dei conigli, per esempio, è interpolata nel tempo da una curva esponenziale. Nel grafico che segue compaiono due curve: quella oscillante rappresenta l'andamento della popolazione dei conigli nel tempo (legge oraria), l'altra è l'esponenziale, sulla quale stanno i punti di massimo.
La crescita delle popolazioni avviene comunque in maniera esponenziale, nel senso che la sequenza dei massimi della popolazione dei conigli, per esempio, è interpolata nel tempo da una curva esponenziale. Nel grafico che segue compaiono due curve: quella oscillante rappresenta l'andamento della popolazione dei conigli nel tempo (legge oraria), l'altra è l'esponenziale, sulla quale stanno i punti di massimo.
Quindi, alla lunga, l'evoluzione delle due specie in conflitto segue lo stesso corso che si avrebbe se fossero separate, obbedendo appunto, pur con alti e bassi, ad una legge di tipo esponenziale.
La funzione logistica
Un altro modello di crescita della popolazione è quello ideato dal matematico e statistico belga P.F. Verhulst (Bruxelles, 28 ottobre 1804 – Bruxelles, 15 febbraio 1849) , che propose di sostituire il fattore crescita costante con un fattore di crescita variabile in funzione del numero di individui della popolazione, assumendo come fattore di crescita la seguente legge di variabilità lineare:
a(x)=-mx+q con m>0
In tale ipotesi il tasso di crescita della popolazione è espresso dalla legge
![]()
cui corrisponde il processo iterativo
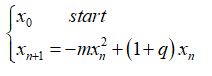
Il modello si basa su una trasformazione non lineare in quanto la variabile x è di secondo grado ed è espresso dalla seguente funzione
![]()
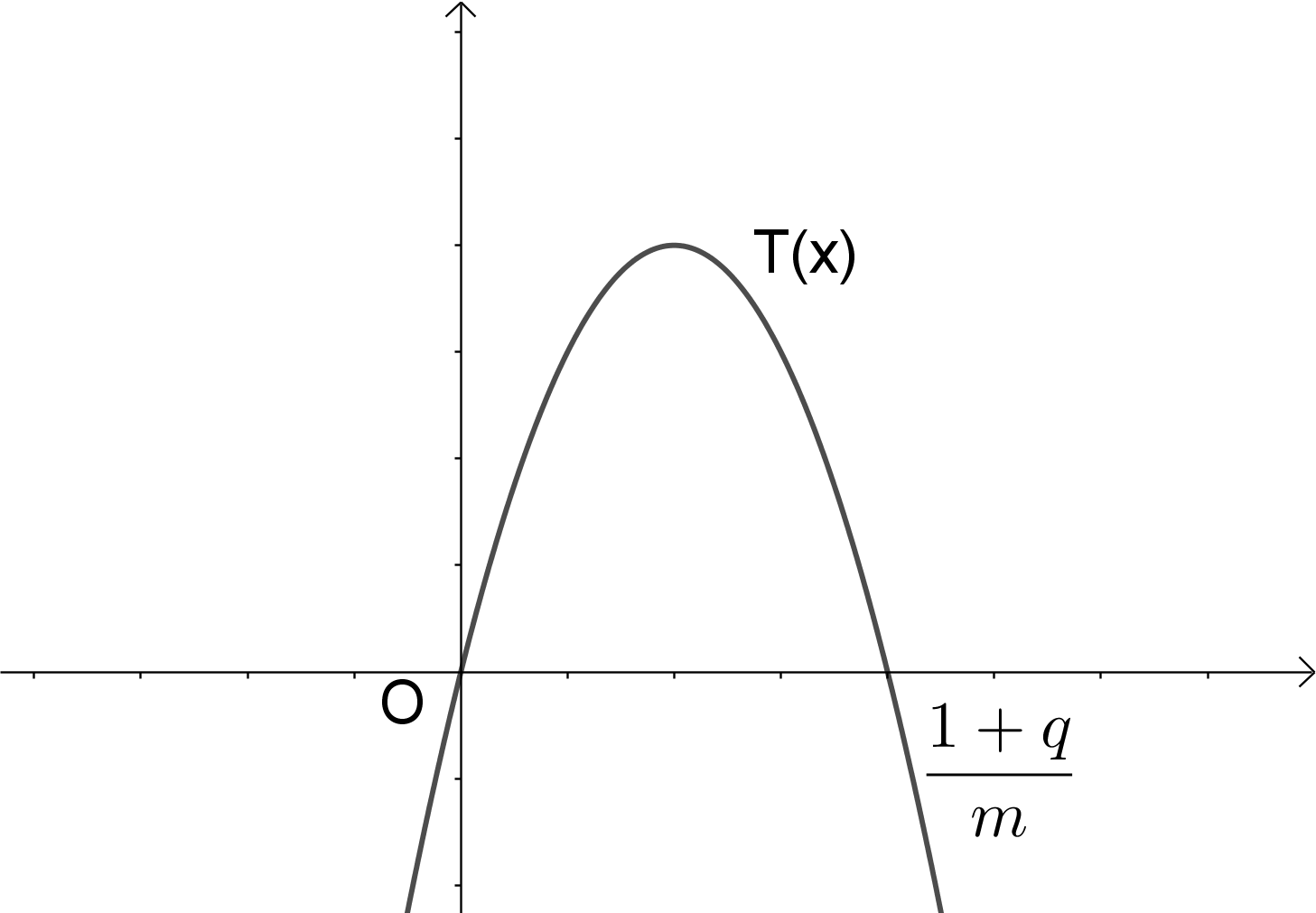
Applicando una trasformazione geometrica (contrazione) alla funzione T(x) è possibile ricondurre il grafico della funzione nell’arco di parabola, delimitato dall’intervallo unitario [0,1]. Se si lascia fissa l’origine degli assi e si pone x=hX con
![]()
il punto
viene trasformato nel punto (1,0) e l’espressione analitica della funzione diventa
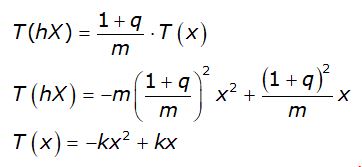
dove si è posto
![]()
di conseguenza il processo iterativo assume la forma
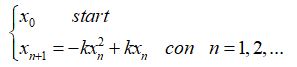
Il nuovo modello differisce da quello di Malthus per il valore
![]()
che rappresenta il termine inibente la crescita.
Le ipotesi su cui si basa questo modello sono: da un lato la crescita della popolazione che si moltiplica in progressione geometrica, dall’altro una resistenza a tale sviluppo direttamente proporzionale al quadrato della popolazione.
Ciclo di crescita di una popolazione cellulare
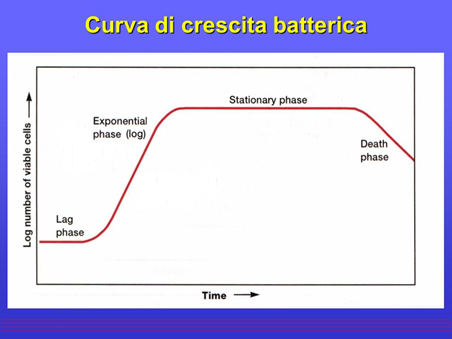 In un contenitore chiuso la crescita esponenziale relativa alla popolazione cellulare è rappresentata da una curva, con andamento quello riportato in figura. Tale curva descrive l'intera dinamica del ciclo di crescita batterica e comprende tre fasi: la fase di latenza, la fase esponenziale (o logaritmica) e la fase di morte.
In un contenitore chiuso la crescita esponenziale relativa alla popolazione cellulare è rappresentata da una curva, con andamento quello riportato in figura. Tale curva descrive l'intera dinamica del ciclo di crescita batterica e comprende tre fasi: la fase di latenza, la fase esponenziale (o logaritmica) e la fase di morte.
La fase latenza
La fase di latenza è il periodo che trascorre tra il momento in cui viene inculcata la popolazione batterica nel terreno e il momento in cui inizia la crescita batterica e può essere più o meno lungo a seconda della provenienza della coltura e delle condizioni di crescita.
La fase esponenziale
Nella fase esponenziale di crescita, ogni cellula si divide per formare due cellule figlie capaci, a loro volta, di duplicarsi per formarne quattro, e così via. Tale fase dura per un periodo breve o lungo in base alle risorse disponibili e ad altri fattori. In tale fase le cellule sono solitamente nel migliore stato di salute e per questo motivo vengono utilizzate per gli studi su enzimi o altri componenti cellulari. Quando la fase esponenziale si blocca per l'esaurimento di un nutriente essenziale del terreno di coltura o per l'accumulo nel terreno di uno o più prodotti del microrganismo fino a livelli inibitori, la crescita entra in fase stazionaria.
La fase stazionaria e di morte
Nella fase stazionaria, il numero delle cellule non aumenta né diminuisce. In alcuni microrganismi si può osservare in questa fase una crescita lenta, ma il loro numero non viene alterato perché alcune cellule si duplicano e altre muoiono (crescita criptica).
Dal momento in cui l'incubazione prosegue oltre la fase stazionaria, le cellule andranno a morire nonostante proseguano, in alcuni casi, il loro metabolismo per un certo periodo. Quest’ultima fase si chiama di morte del ciclo cellulare e in alcuni casi la morte è accompagnata dalla lisi cellulare, cioè dalla demolizione e dissoluzione delle cellule, causata dalla rottura della membrana cellulare. Le diverse fasi della crescita batterica sono eventi che interessano l’intera popolazione cellulare, non la singola cellula.
Questo articolo è sotto la licenza Licenza Creative Commons Attribution Non-commercial No Derivatives 3.0